






研究のおもしろさを! 後輩の諸君へ
同じ道を進もうとしている君への、先輩からの応援メッセージ。
応援のことば
中川 輝良 先生(Department of Chemistry and Biochemistry, University of California, San Diego.)
私は基礎研究を始めたのは、医学部の1年生の時でした。最新の技術を使い独創的な研究をする機会が学生にあるという理由で当時の第一解剖学講座、廣川信隆先生のところで研究を始めました。はじめは分からぬことだらけでしたが、医学部の勉強、所属していた運動会柔道部の稽古に加え、ほとんど毎日夜遅くまで5~12時間位研究室で実験をする事で、自然と手技も身に付き、データも出るようになりました。この結果は、M4のときにJournal of Cell Biologyという当時の細胞生物学のトップジャーナルにfirst authorの論文を掲載することができました1。その過程で学んだ電顕手法は、その後、私の研究において非常に重要な位置を占めています。当時、医学部の教授たちは非常に基礎研究を志す学生に協力的で、しばしば臨床実習を欠席して国内外の学会に自分のデータを発表したりすることも可能でした。ちなみに、学生のとき国際学会で知り合った研究者とは今でも交流があります。
医学部を卒業した後は、 同じ廣川研にて新しいキネシンモーター蛋白の同定と機能解析を行い、PNASとCell誌に発表しました2,3。この結果で学位を取得しました。また、Human Frontier Science ProgramのFellowshipをいただき、2000年より米国ボストンのマサチューセッツ総合病院、Harvard Medical School、HHMIのMorgan Shengの研究室で分子細胞神経科学の分野でのポスドクを行いました。2001年には所属研究室がまるごとマサチューセッツ工科大学(MIT)のPicower Center for Learning and Memoryに移転し、そこで2005年までポスドクをしました。その間、当時、Centerの所長をしていた利根川進先生にはラットのlentiviral transgenesisのプロジェクトを支援して頂きました。さらに、学生のときに習った電顕を再び用い、Harvard Medical SchoolのThomas Walzと共同研究で、今度は構造生物学のプロジェクトを行いました。2005年にNature誌に発表したグルタミン酸受容体の電顕構造の業績により、同年7月カリフォルニア大学サンディエゴ校(UCSD)で独立した研究室を持ち現在に至っています。
最近の研究の詳細については http://nakagawalab.ucsd.edu/ をご覧ください。
いろいろなところを転々として研究を続けてきましたが、今でも東大医学部の学生および大学院時代のトレーニングは非常に重要な基礎となっていると思います。その原点はもちろん学部学生のときに研究を始める機会があり、それにかなりの情熱を注いだところにあります。MD研究者育成プログラムに所属している諸君には『本番の研究人生がすでに始まっている』と意識して頑張ってほしいです。
- Two distinct functions of the carboxyl-terminal tail domain of NF-M upon neurofilament assembly: cross-bridge formation and longitudinal elongation of filaments. Nakagawa T, Chen J, Zhang Z, Kanai Y, Hirokawa N. J Cell Biol. 1995 Apr;129(2):411-29.
- Identification and classification of 16 new kinesin superfamily (KIF) proteins in mouse genome. Nakagawa T, Tanaka Y, Matsuoka E, Kondo S, Okada Y, Noda Y, Kanai Y, Hirokawa N. Proc Natl Acad Sci U S A. 1997 Sep 2;94(18):9654-9.
- A novel motor, KIF13A, transports mannose-6-phosphate receptor to plasma membrane through direct interaction with AP-1 complex. Nakagawa T, Setou M, Seog D, Ogasawara K, Dohmae N, Takio K, Hirokawa N. Cell. 2000 Nov 10;103(4):569-81.
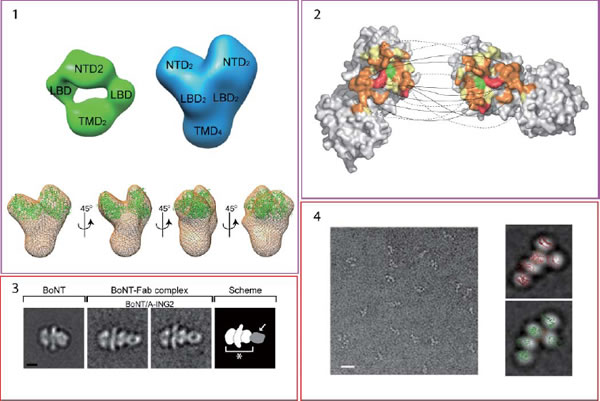
最近の研究で得られた分子構造を簡単に紹介します。
| 1) | AMPA receptor の2量体(緑)と4量体(青)の単粒子電顕構造。4量体電顕構造とN末端ドメイン結晶構造のドッキング像(Nakagawa et al, Nature 2005, Shanks et al, J. Neurosci. 2010、Nakagawa, Mol. Neurobiol. 2010) |
| 2) | NMDA receptor のN末端ドメインの2量体のX-線結晶構造。(Farina et al, J. Neurosci. 2011) |
| 3) | シナプス小包の開放放出を阻害するボツリヌス毒素の単粒子電顕構造。(Fischer et al, 2007, J. Biol. Chem.) |
| 4) | Autismの原因遺伝子の一つであるalpha-neurexinの細胞外ドメインの単粒子電顕構造。(Comoletti et al, Structure 2010) |

学生の皆さんへ
吉田 松生 先生(基礎生物学研究所 生殖細胞研究部門)
私は、小さい頃から昆虫が好きで生き物に興味を持っていました。M3の時には萩原一フェローとしてカリフォルニアの研究所で一夏を過ごし、研究の楽しさを知りました。卒業する時には、臨床と研究の間で迷いました。臨床は、今まで人類が蓄積して来た知識と技術を、目の前の患者さんに的確に提供する仕事です。一方、生物学研究は、人類が知らない生き物の姿を自身の興味に従って明らかにする営みです。私にとって、この2つを両立するのは難しく、どちらかを諦めると先に決めて、研究を選びました。皆さんの多くが持っている「治せない病気を治すために研究しよう」という使命感が希薄だっただけのことですが、お恥ずかしい事にそういう学生でした。
私は基礎生物学の世界に身を置いています。一番大切なのは、「何を問うか?」です。これは、自分で決めなければなりません。ある時、子供に遺伝情報が正しく伝わる秘密を知りたいと思い、マウスの精子幹細胞研究を始めました。既成の手法はピンと来なかったので、自分の思うように研究しようと思いました。それまで何も成し遂げていませんでしたが、やせ我慢を理解してもらえたメンターに感謝しています。7年かかって、幹細胞(と言われている細胞)の動く姿を初めて目にした時は、とても嬉しかったです。一気に世界中の研究者と友達になりました。人と人をつないでくれるサイエンスのすばらしさを実感しました。メンターにも少しだけ恩返しできたかなと思っています。最近、本当に使える応用研究と本当に重要な基礎研究は同じ所に収束することに気づき、学生の時からの悩みも少しずつ解け始めています。しかし、まだまだ道のりは遠く、生き物の奥深さに悪戦苦闘の日々です。
MD研究者育成プログラムは、臨床と研究の間で揺れる学生さんの事を良く考えてあり、短期間に多くの経験を積みやすくなっていると思います。私が学生だったら参加したいと思ったことでしょう。もっとも、参加したからといって、悩むことなく自然に立派な医学研究者になれた、ということもなかったでしょうが。(ユートピアはどこにもないという意味ですよ。誤解のなきよう。)学生の皆さんは、何も成し遂げていない分、あらゆる可能性があります。恵まれた環境を最大限活かして、人生を切り開いてください。
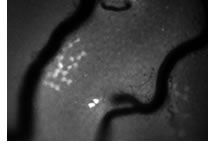 蛍光タンパク質を用いて、生きた精巣中で初めて可視化された未分化型精原細胞。この細胞群のライブイメージングによって幹細胞の実体を少しずつ明らかにしている。詳しくは:http://www.nibb.ac.jp/germcell/
蛍光タンパク質を用いて、生きた精巣中で初めて可視化された未分化型精原細胞。この細胞群のライブイメージングによって幹細胞の実体を少しずつ明らかにしている。詳しくは:http://www.nibb.ac.jp/germcell/
【略歴】1991年東京大学医学部医学科卒、 1995年医科学研究所にて博士課程修了(新井賢一先生)。国立精神・神経センター研究員、大阪大学細胞生体工学センター助手、京都大学医学研究科助教(鍋島陽一先生)を経て、2008年より自然科学研究機構基礎生物学研究所教授。
チャンスをつかめる技術を身につける
粂 和彦 先生(熊本大学発生医学研究所 )
ぼくはM1のFQで養老孟司先生の解剖学教室に、M2は脊山洋右先生の栄養学教室に入りました。そこで当時助教授の清水孝雄先生と出会い、夜と週末は研究室に入り浸り、さらにM4の夏休みをNIHにいた岡山博人先生のところで過ごしました。渡航費は内科の勉強会に参加させてもらっていた高久史麿先生が出してくれました。卒業時に、ぼくは生意気だったので、科学者としては最も進んだ最先端の研究を、医師としては最も患者さんに近い最前線の医療の両方をしたいと考えました。当時の大学病院の医療や研修は自分の理想からは遠かったため、最前線でのオールラウンドの力を短期間でつけられる民間病院のローテート研修を2年間受けました。その後、帰国された岡山先生の大学院に入り、博士課程修了後は清水先生の助手にして頂き、ずっと分子生物学の研究を続け、留学を契機に、概日周期と睡眠覚醒制御をメインテーマにしています。一方、医師としての仕事も大学とは独立に独力で続けてきた中で、熊本県には3名しかいない睡眠医療の認定医となり、睡眠障害診療でも一定の役割を果たしています。卒後20年して初めて、医師としての仕事と研究者としての仕事が、「睡眠」という同じキーワードでつながりました。とは言っても、今でも若い頃の理想どおり、研究者としては純粋に科学的な興味を追及し、医師としては、あくまでプライマリーケアにこだわっていますが、難しい症例であればあるほど、自分の科学的な経験・視点も役に立っています。良い「医療」のためには、知識としての医学だけではなく、科学としての医学の視点がまちがいなく重要で、そのような視点は、自分自身が「科学」の中で未知のものと対決しない限りは、本当の意味では身につかないと思います。今回、始まるMD研究者育成プログラムは、ぼく自身の学生時代の経験をシステム化したように思えて、東大の学生さんたちを羨ましく感じるほどです。是非、活用して下さい。
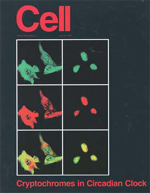 |
Cell誌の表紙を飾った写真。クリプトクロームというタンパク質が、時計遺伝子のピリオドのタンパク質を核内に移行させることを、世界で初めて示した写真です。画期的な結果だったので、撮った時には本当に興奮しました。この成果は留学後3ヶ月という短期間で発表できたので、もちろん運が良かったのですが、個々の実験を失敗なく最短時間で終えることができる確固とした技術を身につけていたことが重要だったという自負もあります。 |
粂先生の研究の詳細は:http://k-net.org/
「とりあえず」でなく「やるべきこと」を!
加藤 忠史 先生(理研脳科学総合研究センター 精神疾患動態研究チーム)
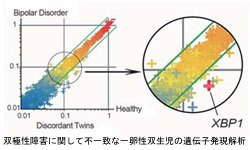 双極性障害に関して不一致な 一卵性双生児の遺伝子発現解析 |

15名の国際的研究者よりなる外部評価チームを前に、5年間のチームの研究成果を発表
加藤先生の研究の詳細は:
http://www.brain.riken.go.jp/labs/mdmd/


 HOME
HOME